The previous two sections showed how the afferent and lateral connections self-organize into a highly structured map with very specific internal connections, as found in the cortex. As one might expect from the properties of the individual neurons in the map, the response of the network to an input varies systematically depending upon the orientation and position of that input. This section will show examples of the network's actual response to different orientations, and will examine possible methods for determining what orientation is perceived by the cortex for that activity pattern. Calculating the perceived orientation is an essential prerequisite to measuring tilt aftereffects and illusions, since those are manifested as differences in the perceived orientation in different circumstances.
At any point in time the visual system is processing an image that has
only a small number of oriented features in any local area, so only a
few portions of the orientation map will be active at a given time.
Figure 4.6 shows the sparse activity
that results for Gaussian inputs of various orientations at the center
of the retina.
![\begin{figure}
\begin{minipage}
{\textwidth}
\centering
\begin{minipage}
{\thr...
...\makebox[0pt][c]{
\usebox {\orkey}
}
\end{minipage} \end{minipage} \end{figure}](img55.gif) |
However, it is unknown precisely how the higher visual areas extract the encoding of orientation from figure 4.6 and arrive at the perception of an individual oriented line. It has been suggested that the perceived orientation is either the orientation preference of the unit with the highest activation (Carpenter and Blakemore, 1973), or a weighted average of the preferences of all active units (Coltheart, 1971). An intermediate method could also be used, computing an average of all the units having activity greater than some arbitrary activity level. In order to determine if a choice between these methods is crucial, the two extreme options were tested for the RF-LISSOM model.
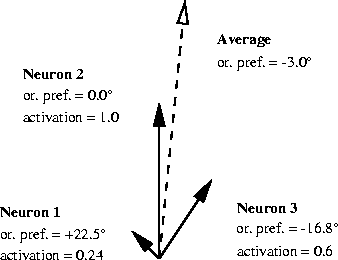
The weighted average of orientation preferences must be computed as a vector sum, since angles repeat every 180°. Two nearly horizontal lines (e.g. -85° and +85°) should average to represent a horizontal line (±90°). However, the arithmetic average of -85° and +85° is 0°, which is a vertical line and is clearly incorrect as an estimated perception. Instead, each neuron is represented by a vector. The vector must represent adjacent orientations as adjacent vector angles. Thus each of the 180° possible orientations must be scaled by two to get the angle of the vector, which ranges over 360°. Since each neuron is to contribute only to the extent that it is active, the magnitude of the vector is taken to be the activation level of the neuron. Once the neurons have been represented in this fashion, the average orientation can be computed from the orientation of the vector sum. Figure 4.7 illustrates these calculations.
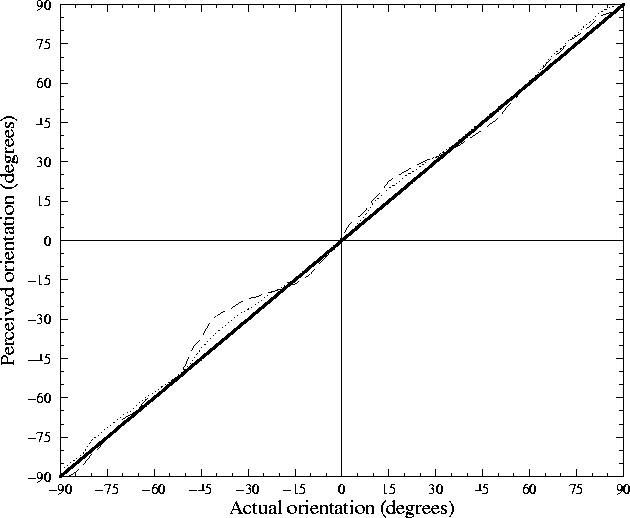
Using the vector sum method of computing the average, the perceived orientation was calculated for the trained network using both the average value and the maximum value methods. For each each angle, an oriented Gaussian (of the same shape as in training) was presented at the center of the retina, and the estimated orientation was computed using each method. The results are presented in figure 4.8.
Both methods appear to be reasonably accurate representations of orientation. Small deviations from the true angle are present, however, because the activated cortical area is comparable to the size of an orientation column. The number of neurons with RFs receptive to the peak of the input Gaussian is relatively small, and thus the distribution of orientation preferences will not necessarily be uniform within that population. Thus, at different locations on the retina, the estimated orientation will differ slightly. If much larger inputs are used, activating a large cortical area, these effects should cancel out, allowing arbitrarily accurate orientation encoding. However, this would be computationally prohibitive to verify at present since much larger cortex and retina sizes would need to be simulated.
As one might expect, utilizing all active units in the calculation of the perceived orientation is slightly more accurate than only using those at maximum activation, but the difference is not significant because many neurons are at maximum and thus most of the neurons averaged in each case are identical. For simplicity, I have chosen in later experiments to use only the method of averaging all units, but the results should be the same for either method, or for any similar method. Note that there is insufficient biological evidence to support a particular method, and I do not claim that orientation perception actually need be occurring in precisely this way. In any case, to ensure that the biases shown in figure 4.8 do not distort the results, all perceived orientation measurements in this thesis are stated in terms of differences in perceived angles, rather than in terms of the actual orientation on the retina.