


In this section we want to analyze, in quantitative form, how the resultant RF depends on the presence and the values of various architectural primitives. Specifically we investigate the influence of the orientation map and of the inhibitory schema.
The order of magnitude specification of the parameters characterizing the model, can be guided by physiological findings:
 , as it occurs for RF centers located between
, as it occurs for RF centers located between  and
and
 degrees from the fovea
[2].
degrees from the fovea
[2].
 ) in order that the average diameter of the resulting RF is
approximately twice the average wavelength (
) in order that the average diameter of the resulting RF is
approximately twice the average wavelength ( ) of the orientation
map. This ensures that the RFs of cells separated by a distance of
) of the orientation
map. This ensures that the RFs of cells separated by a distance of
 do not overlap, as reported by numerous experimental
observations
[2,58,60,61,107].
do not overlap, as reported by numerous experimental
observations
[2,58,60,61,107].  and their relative displacement at
and their relative displacement at
 , while the excitatory gain a is kept at
, while the excitatory gain a is kept at  [64,65,67,123,125].
[64,65,67,123,125].
 are shown
in the insets.
are shown
in the insets.
 |
 |
 |
 |
 |
 (a and b) and
(a and b) and  (c and d),
respectively.
The insets show the behavior of the equivalent feed-forward kernels is shown,
for different values of the inhibition strength b (
(c and d),
respectively.
The insets show the behavior of the equivalent feed-forward kernels is shown,
for different values of the inhibition strength b ( to
to  , by
step of
, by
step of  ).
).
For low values of b, in addition to the dominant central subregion,
supplied by LGN projections, we observed in the resulting RF the presence
of parallel inhibitory flanks, which prevent responses to non-preferred
orientations. Increasing the strength of inhibition, additional sidebands
appear in the RF, endowing it with a well-defined multipartite structure
that resembles that of real ``periodic'' simple cells
[29,88,120].
The clustered nature of the modeled inhibition manages, indeed, to
establish spatial periodic induced couplings with surrounding cells that are
alternately positive and negative.
Specifically, positive couplings take place between cells with overlapping
inhibitory fields. Such induced excitatory fields contribute,
in turn, to the formation of additional negative or positive couplings with
other cells, depending on the signs of their overlapping interaction
fields. Thus, recurrent inhibition results in
an increasing extension of effective coupling over cortical plane as it
appears well evidenced in the shape of the equivalent feed-forward kernel
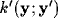 (see figure 4).
(see figure 4).