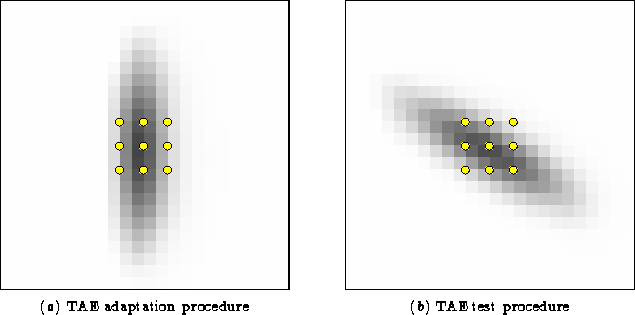
![[*]](foot_motif.gif) The state of the map at 30,000 iterations
was taken as a starting point for each independent experiment in this
chapter, so the results roughly correspond to testing a single human
subject under different conditions. To simulate adapting to an
oriented stimulus as in the psychophysical experiments, the position
and angle of the inputs were fixed to a single value for a number of
iterations (figure 5.1a).
The state of the map at 30,000 iterations
was taken as a starting point for each independent experiment in this
chapter, so the results roughly correspond to testing a single human
subject under different conditions. To simulate adapting to an
oriented stimulus as in the psychophysical experiments, the position
and angle of the inputs were fixed to a single value for a number of
iterations (figure 5.1a).
As adaptation progressed, test lines at various orientations were presented without modifying any weights (figure 5.1b). For each test line, the perceived orientation was measured as described in section 4.5. The magnitude of the tilt aftereffect was defined as the perceived orientation of the test line after adaptation minus the perceived orientation before adaptation.
This procedure is similar to the procedure used for human subjects described in section 2.2.1. For computational efficiency, it takes advantage of the fact that learning can be turned off in the model. Since experiments with humans cannot turn off learning for testing, they cannot evaluate the effects of a single adaptation episode on multiple test orientations equally. That is, the data for the second test figure presented after adaptation would be affected by the presentation of the first test figure. Instead, psychophysical experiments use a single test figure. They evaluate the effect on its perceived orientation by adaptation at different angles relative to the test figure (see section 2.2.1). This alternative procedure is computationally prohibitive to simulate (not to mention extraordinarily time-consuming to measure in humans) because one must ensure that the cortex has returned to equilibrium between the adaptation episodes. Since the angular function of the tilt aftereffect has been demonstrated to be similar for all orientations (Mitchell and Muir, 1976), the two procedures should give an equivalent measure of the amount of aftereffect at each angle. The procedure used here allows much more comprehensive data to be collected for the model, as shown in section 5.5.
Because the TAE curves differ substantially between individuals, particularly in the zero crossing between direct and indirect effects, it can be misleading to average results from different subjects or testing paradigms. For instance, if the zero crossings vary over some range, a null area will show up in the graph around that region, even though no individual exhibited a null area. However, since the data are generally too erratic to interpret from a single run, multiple runs from a single individual are usually averaged (e.g. Mitchell and Muir 1976).
To obtain similar measurements for the RF-LISSOM model, a single orientation map was tested separately at 9 different positions forming a 3 × 3 grid covering a retinal area 6 × 6 units around the center of the retina (figure 5.1). This range of positions covers many different orientation preferences. Averaging over these positions reduces the random fluctuations in TAE magnitude (see section 4.5), but it does not change the basic shape of the curve presented in the following section.